


Россия
УДК 59 Зоология
УДК 62 Инженерное дело. Техника в целом. Транспорт
Проведен анализ научных публикаций по эхолокации морских млекопитающих, в которых приводятся значения частот следования зондирующих импульсов, излучаемых морскими животными, существенно превышающих частоты следования, необходимые для определения расстояния до объектов охоты. Установлена взаимосвязь между повышенными частотами следования импульсов в сериях и типами объектов охоты. Частоты следования импульсов данного типа и соответствующие им объекты охоты сведены в таблицу.
морские млекопитающие, эхолокация, частота следования импульсов
Введение
С учетом протяженности морских границ, важнейшей компонентой национальной безопасности России является Военно-морской флот. За последнее десятилетие США и страны Запада разработали большое количество подводных роботов различных классов, изготовленных из современных материалов. Вероятность их запуска с атомных подводных лодок в территориальные воды России достаточно высокая. В связи с этим необходима доработка алгоритмов гидроакустических станций надводных кораблей и подводных лодок для обнаружения и классификации подводных объектов, изготовленных из различных материалов. Существующие алгоритмы работы гидроакустических станций не позволяют определять материал, из которого состоит тот или иной объект локации, в то же время известно, что зубатые киты обнаруживают и классифицируют моллюсков и рыб на значительных расстояниях.
В данной работе проведен анализ научных публикаций по эхолокации морских млекопитающих, в которых приводятся значения частот следования зондирующих импульсов, излучаемых морскими животными, существенно превышающих частоты следования, необходимые для определения расстояния до объектов охоты.
В настоящее время существует несколько гипотез, объясняющих причину излучения морскими млекопитающими зондирующих импульсов с повышенными частотами следования внутри пакетов (сериях). Суть гипотез сводится к тому, что при приеме нескольких эхосигналов от мишени увеличивается отношение сигнал/помеха [1].
Анализ существующих публикаций и новых экспериментальных данных позволяет высказать еще одну гипотезу о причине выбора повышенной частоты следования импульсов.
Известно, что интенсивность отраженного сигнала зависит от акустического сопротивления мишени, т.е. произведения плотности материала мишени и скорости распространения звука в мишени. В тех случаях, когда акустическое сопротивление мишени близко к акустическому сопротивлению воды, интенсивность отраженного сигнала мала. Вероятно, что в этих условиях обнаружение моллюсков и рыб морскими млекопитающими происходит за счет переизлучения от них на собственных резонансных частотах. Данный эффект возникает при излучении зондирующих импульсов, согласованных с типом и размером моллюсков, рыб. Для обнаружения подводных объектов на больших расстояниях и в условиях помех зондирующие импульсы морских млекопитающих следуют внутри пакетов с частотой, согласованной со свойствами материала объекта поиска. Возможно, что в этом случае эффект переизлучения от объектов поиска усиливается.
Анализ научных публикаций по эхолокации морских млекопитающих
Исследование возможностей биосонаров китообразных проведено в работе [2]. Мишени (кальмары) размещались под различными ракурсами относительно направления излучения смоделированных зондирующих импульсов зубатых китов Physeter macrocephalus, Mesoplodon densirostris, Orcinus orca L. и Tursiops truncatus. Отмечено, что полученные дальности обнаружения мишени превышают дальности, рассчитанные по уравнению гидролокации с учётом отражающей способности цели и времени интегрирования эхосигнала (приблизительно 264 мкс). Из статьи [2] следует, что существующие гидроакустические станции, обладающие достаточной энергетикой, не могут обеспечить получение таких же дальностей обнаружения кальмара, какие могут быть получены с помощью биосонаров зубатых китов.
В работе [3] проведено сравнение частот следования эхолокационных импульсов косаток Orcinus orca L. в природе при разных формах активности (охоте и перемещении). Приведены параметры (медианы) эхолокационных серий косаток при разных формах активности:
Форма активности Частота следования импульсов, Гц
Охота на терпуга 11.26
Охота на лосося 14.86
Перемещение 7.20
Также приводятся зондирующие импульсы косаток с повышенной частотой следования при разных формах активности:
Форма активности Частота следования импульсов, Гц
Охота на терпуга 205.11 ± 5.90
Охота на лосося 160.80 ± 7.60
Перемещение 203.60 ± 9.60
Авторы статьи [3] отмечают, что лосось крупнее терпуга и энергетически более выгоден с точки зрения питания, однако он значительно более быстр и ловок, и на его поимку требуется много времени и энергии.
Исходя из приведенных данных, и если допустить, что между повышенными частотами следования импульсов внутри пакетов и типами объектов охоты есть взаимосвязь, можно предположить, что косатки, излучающие зондирующие импульсы при перемещении с частотой следования 203.6 Гц, на самом деле продолжают искать терпуга (частота 205.11 Гц). Вероятно, что для косаток терпуг является более предпочтительным объектом охоты, чем лосось.
Частоты следования импульсов 150–210 Гц не связаны с акустическими полями рыб, рассмотренными в [4]. Они не являются сигналами, которые китообразные могли бы излучать для приманивания рыб, т.к. такие же частоты следования импульсов использовал дельфин, которому в качестве мишени предъявлялся кусок рыбы [5, 6]. Опыт заключался в том, что дельфин-бутылконос Tursiops truncatus с присосками на глазах и находящийся в круглом бассейне диаметром 10.5 м и глубиной 1.65 м по сигналу исследователей направлялся к рычагу звонка, опущенному в воду, и нажимал на него, в результате чего звонил звонок, затем плыл к исследователю за наградой. В качестве награды в воду (на расстоянии 2.5–5 см перед гидрофоном) опускался кусок рыбы (макрель) с поперечным сечением около 5 см. Когда дельфин приступал к выполнению задания, он начинал издавать поисковые звуки в виде импульсов с частотой повторения около 16 Гц. За время, пока дельфин подходил к объекту поиска, частота повторения его звуковых импульсов возрастала и достигала 190 Гц в тот момент, когда дельфин брал кусок рыбы. Затем частота повторения падала до 26–50 Гц. Отмечено, что дельфин не касался гидрофона, когда брал кусок рыбы.
Экспериментаторы считали объектом поиска кусок рыбы, а “ослеплённый” дельфин, из соображений безопасности, вынужден был учитывать, что в бассейне кроме куска рыбы находятся ещё три объекта: гидрофон (стальной корпус), шнур гидрофона (из полихлорвинила) и рычаг.
Дельфин решал задачу поиска куска рыбы в условиях помехи - гидрофона. Расстояние до гидрофона и до куска рыбы было одинаковое.
Предположим, что сначала дельфин, используя частоту повторения импульсов 16 Гц, обнаружил гидрофон, а затем, двигаясь в сторону гидрофона и значительно увеличив частоту повторения импульсов, обнаружил кусок рыбы перед гидрофоном. После этого дельфин использовал частоту 26–50 Гц для поиска рычага или другого ориентира для возвращения в исходную точку.
Основанием для такого предположения является анализ записей локационных сигналов белухи Delphinapterus leucas (Pallas, 1776) из массива записей, описанных в [7], где частота следования импульсов белухи по стальному цилиндру была ~16 Гц. Это видно из рисунка 1, где на представленном фрагменте записи локационного сигнала белухи длительностью 0.4 с определяются 6 зондирующих импульсов.
Материалы и методы
Для анализа структуры сигналов белухи Delphinapterus leucas, представленных на рисунках 1–4, использованы записи сигналов, собранные в 2018 году авторами статьи [7]. При проведении опытов белуха находилась в вольере. Мишени размещались за пределами вольера на различных расстояниях. Частота дискретизации в файлах с записями сигналов белухи – 1 МГц. Запись производилась с гидрофонов B&K 8103. Расстояние между гидрофонами 2 м.
Скорость звука в соленой воде во всех экспериментах ~ 1440 м/с.
Фрагменты записей локационных сигналов белухи выбирались из файлов, где белуха определила тип мишени правильно.
Частоты следования импульсов внутри пакетов определялись только по тем зондирующим импульсам, под действием которых переизлучение от мишени было максимальным.
Уровни сигналов на рисунках указаны в безразмерных величинах.
Анализ параметров локационных серий белухи
На рисунке 1 приведён фрагмент записи локационного сигнала белухи по мишени типа “стальной цилиндр”. Расстояние до мишени 60 м. Частота следования импульсов в середине серии изменялась в пределах: 16–16.3 Гц. Расчетная дальность без коэффициента перекрытия ~45 м.
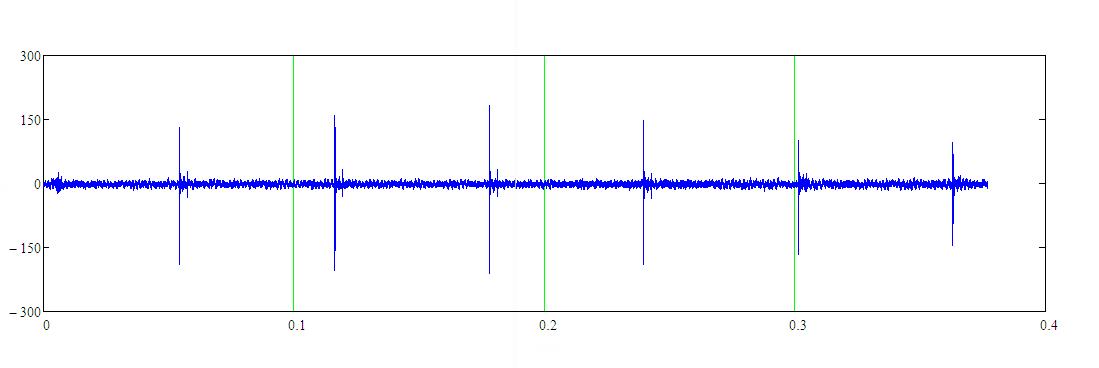
Рисунок 1 – Фрагмент записи локационного сигнала белухи
по мишени типа “стальной цилиндр”
На рисунке 2 приведён фрагмент записи локационного сигнала белухи длительностью 0.35 с по мишени типа “стальной цилиндр” (с двух гидрофонов). Расстояние до мишени 43 м. Частота следования импульсов в середине серии изменялась в пределах: 24–24.5 Гц. Расчетная дальность без коэффициента перекрытия ~30 м.
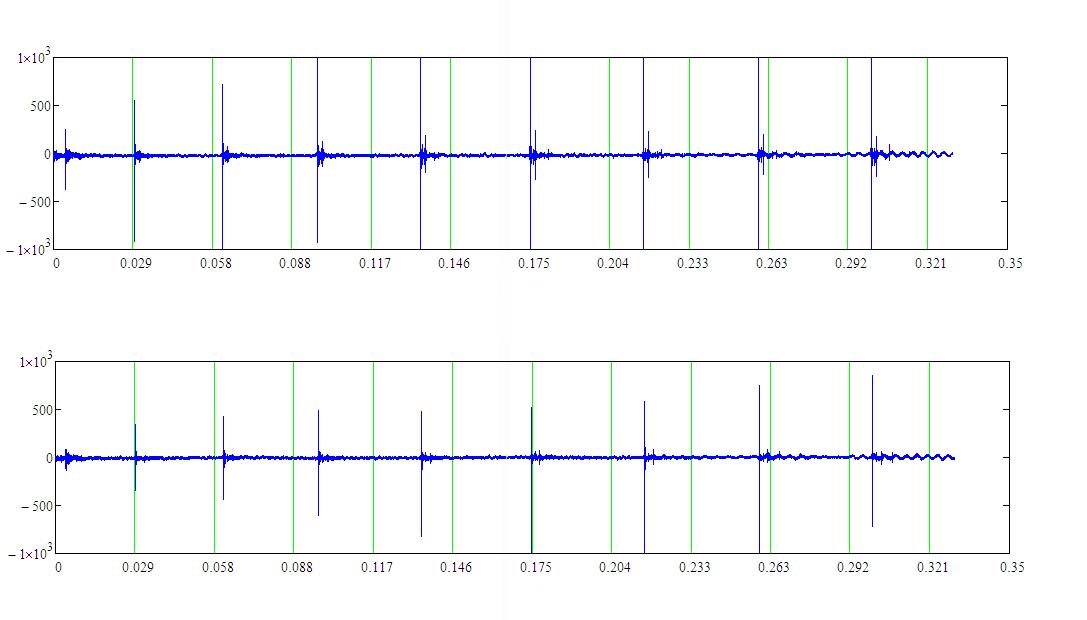
Рисунок 2 – Фрагмент записи локационного сигнала белухи по мишени типа
“стальной цилиндр” с левого (вверху) и правого (внизу) гидрофона
На рисунке 3 приведён фрагмент записи локационного сигнала белухи длительностью 0.4 с по мишени типа “стальной шар”. Расстояние до мишени 60 м. Частота следования импульсов в середине серии равна 8.4 Гц. Расчетная дальность без коэффициента перекрытия ~85.7 м.
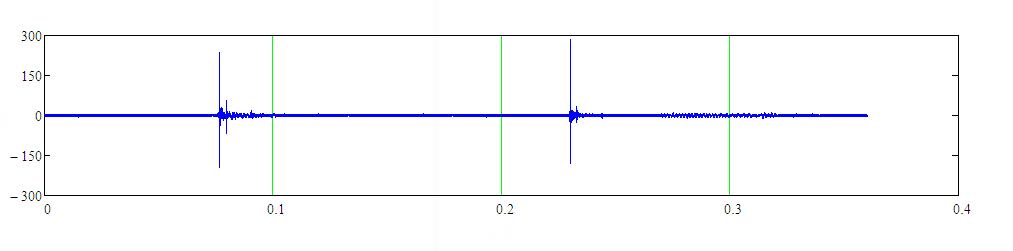
Рисунок 3 – Фрагмент записи локационного сигнала белухи
по мишени типа “стальной шар”
На рисунке 4 приведён фрагмент записи локационного сигнала белухи длительностью 0.18 с по мишени типа “полистироловая крышка” (с двух гидрофонов). Расстояние до мишени 15 м. Частота следования импульсов в серии изменялась в пределах: 28.2-30.64 Гц. Расчетная дальность без коэффициента перекрытия ~24 м.
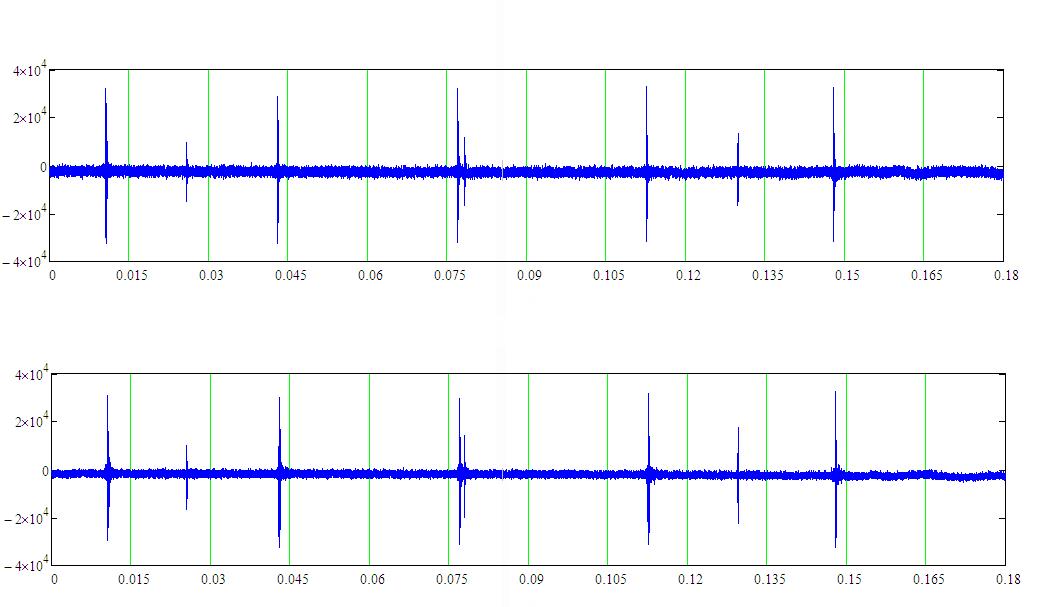
Рисунок 4 – Фрагмент записи локационного сигнала белухи по мишени типа “полистироловая крышка” с левого (вверху) и правого (внизу) гидрофона
Результаты
Сведём полученные повышенные частоты следования зондирующих импульсов морских млекопитающих в таблицу 1.
Таблица 1.
Частоты следования зондирующих импульсов морских млекопитающих
в зависимости от объекта поиска
|
Объект поиска |
Частота, Гц |
Примечание |
|
Рыба терпуг |
205 |
из [3] |
|
Рыба макрель (кусок) |
190 |
из [5] |
|
Рыба лосось |
160 |
из [3] |
|
Крышка (полистирол) |
30 |
из рисунка 4 |
|
Цилиндр (сталь) |
16 и 24 |
из рисунков 1-2 |
|
Шар (сталь) |
8 |
из рисунка 3 |
Следует обратить внимание, что повышенные частоты следования зондирующих импульсов морских млекопитающих по стальным мишеням кратны 8 (8, 16, 24).
Обсуждение результатов
Данные таблицы 1 позволяют установить, что для мишеней (рыб) частоты следования импульсов на порядок отличаются от частот в сериях, излучаемых морскими млекопитающими по мишеням из стали и полистирола.
На рисунке 2 в конце фрагмента записи (временной участок 0.263–0.321 с) видно переизлучение от стального цилиндра на одной из собственных резонансных частот.
Резонансное переизлучение возникает только в том случае, когда форма и длительность зондирующих импульсов согласована с параметрами мишени. Известно, что скорость распространения звуковых колебаний в материалах мишеней различна для разных типов мишеней. В работе [8, с.425] отмечено, что “…несмотря на трехкратное различие в волновых сопротивлениях, мишени из стали и дюраля совершенно не различаются даже при минимальном удалении животного от них. Таким образом, именно скорость сдвиговых волн в материале, а не акустическое сопротивление последнего позволяет дельфинам различать материал мишеней”.
Так как в тканях моллюсков и рыб не могут распространяться сдвиговые волны (поперечные колебания), а они, тем не менее, обнаруживаются морскими млекопитающими на значительных расстояниях, то остается предположить, что зондирующие импульсы внутри пакетов, следующие с определенной частотой, возбуждают поверхностные волны в материале мишени. Возможно, что согласованные с материалом мишени пакеты зондирующих импульсов могут не только препятствовать затуханию поверхностных волн, но и усиливать их пульсации.
Данное предположение не противоречит выводам исследователей, проводивших анализ структуры эхосигналов от упругих шаров:
– о возможности возбуждения и распространения в материале мишени поверхностных волн;
– об использовании дельфином информации о времени прохождении зондирующего сигнала через мишень в качестве информативного признака для распознавания.
Эти выводы приведены в книге [9, с.83].
Табличные значения скоростей распространения поверхностных волн в справочниках найти трудно, но данные о скорости распространения поперечных колебаний (для твердых тел) в справочной литературе широко представлены. Их значения на 7-8% больше скоростей распространения поверхностных волн Рэлея. Этого достаточно для приблизительного оценивания скорости распространения поверхностных волн на металлических цилиндрах по скорости поперечных колебаний.
Для набора статистики о соответствии между частотами следования импульсов и типами объектов охоты необходимо проведение натурного эксперимента с мишенями, имеющими различные акустические сопротивления и скорости распространения поверхностных волн.
Натурный эксперимент должен быть проведен по определённой схеме. Обученным морским млекопитающим различных видов должны предъявляться два типа мишеней: рыба (кусок рыбы), диаметром не менее 5 см и прямой круговой металлический цилиндр (без полостей), диаметром не менее 5 см. Для мишеней - рыб необходимо делать запись об объеме плавательного пузыря или об его отсутствии. При проведении экспериментов в бассейне морским млекопитающим для чистоты эксперимента необходимо надевать присоски на глаза. Расстояние до мишени должно выбираться таким, чтобы был исключен с ней визуальный контакт. После предъявления в случайном порядке двух типов мишеней статистически значимое число раз и обнаружения существенной разницы частот следования зондирующих импульсов внутри пакетов для рыб и для цилиндров, при условии правильного распознавания, делается вывод о подтверждении гипотезы резонансного механизма переизлучения от мишеней за счет согласованных с ними частот следования зондирующих импульсов.
Выводы
1. Установлена взаимосвязь между повышенной частотой следования зондирующих импульсов морских млекопитающих и типами объектов охоты.
2. Необходимо проведение натурного эксперимента с мишенями, имеющими различные акустические сопротивления и скорости распространения поверхностных волн с целью дополнить таблицу соответствия (таблица 1), что позволит получить дополнительную информацию о типе объекта охоты, который заинтересовал морское животное, путём анализа повышенных частот следования импульсов морских млекопитающих и условий их записи.
3. Для обнаружения подводных объектов на больших расстояниях и в условиях помех зондирующие импульсы перспективных гидроакустических станций должны излучаться пакетами. Частота следования импульсов внутри пакетов должна быть согласована со скоростью распространения поверхностных волн в материале объекта поиска.
4. Данный подход позволяет проводить ретроспективный анализ публикаций по эхолокации морских млекопитающих. Например, из книги “Гидролокатор дельфина” [9, с.19] можно узнать, что “существенная разница в частоте следования импульсов в сериях при дифференцировке животными мишеней позволяет разделить их на серии с малой частотой следования 20-40 Гц и серии с большой частотой следования 100-400 Гц. Серии с большой частотой следования характерны для начальных опытов, когда только вырабатывается условный рефлекс у животного. Можно предположить, что такие серии используются животными с целью ориентации”.
Исходя из того, что между повышенными частотами следования импульсов внутри пакетов и типами объектов охоты установлена взаимосвязь, существенное различие частот следования импульсов в сериях, обнаруженных в [9], можно объяснить тем, что на первом этапе обучения дельфину в бассейн бросали рыбу, а на втором этапе в бассейне размещали мишени из различных материалов для их дифференцировки.
1. Иванов М.П. Эхолокационные сигналы дельфина (Tursiops truncatus) при обнаружении и распознавании подводных объектов, дис. …канд.биол. наук, СПб., 2000. - 137 с.
2. Madsen P.T., Wilson M., Johnson M., Hanlon R.T., Bocconcelli A., Aguilar de Soto N., Tyack P.L. Clicking for calamari: toothed whales can echolocate squid Loligo pealeii // Aquatic biology, 2007, Vol. 1. - P. 141-150.
3. Лазарева Е.М., Филатова О.А., Бурдин А.М., Хойт Э. Параметры эхолокационных серий косаток (orcinus orca) при различных формах активности // Зоол. журн. - 2010. - Том 89. - № 5. - С. 625-631.
4. Электрические и акустические поля рыб / АН СССР. Институт эволюционной морфологии и экологии животных им. А.Н. Северцова. - М.: Наука, 1973. - 232 с.
5. Norris K.S., Prescott J.H., Asa-Dоrian P.V., Perkins P. An experimental demonstration of echolocation behaviour in the porpoise, Tursiops truncatus (Montagu). Biol. Bull., 1961, 120, 2. - P. 163-176.
6. Романенко Е.В. О гидролокационной способности дельфинов обзор // Акуст. журн., 1964, том Х, № 4. - С. 384-397.
7. Цветков О.В., Сиренко А.И., Мухачёв Е.В., Данилов Н.А., Исаков Д.Ю., Овчинников К.Э., Розум А.В., Инякина Н.В., Лазуренко Т.В., Сидоров Р.А., Соколов П.А. Теоретические и экспериментальные исследования бионических принципов гидролокации // Прикладные проблемы безопасности технических и биотехнических систем. - 2018. - № 2. - С. 2-13.
8. Айрапетянц Э.Ш., Константинов А.И. Эхолокация в природе / АН СССР. Отделение физиологии. - 2-е изд., перераб. и доп. - Л.: Наука, 1974. - 512 с.
9. Голубков А.Г. Гидролокатор дельфина. - Л.: Судостроение, 1977. - 96 с.